1.1 Elements of Neuronal Systems
Over the past hundred years, biological research has accumulated an enormous amount of detailed knowledge about the structure and function of the brain. The elementary processing units in the central nervous system are neurons, which are connected to each other in an intricate pattern. A tiny portion of such a network of neurons is sketched in Fig. 1.1, which shows a drawing by Ramón y Cajal, one of the pioneers of neuroscience around 1900. We can distinguish several neurons with triangular or circular cell bodies and long wire-like extensions. This picture can only give a glimpse of the network of neurons in the cortex. In reality, cortical neurons and their connections are packed into a dense network with more than cell bodies and several kilometers of ‘wires’ per cubic millimeter. Across areas of the brain the wiring pattern may look different. In all areas, however, neurons of different sizes and shapes form the basic elements.
Still, the cortex does not consist exclusively of neurons. Beside the various types of neuron, there is a large number of ‘supporter’ cells, so-called glia cells, that are required for energy supply and structural stabilization of brain tissue. Since glia cells are not directly involved in information processing, we will not discuss them any further. We will also neglect a few rare subtypes of neuron, such as non-spiking neurons in the mammalian retina. Throughout this book we concentrate on spiking neurons only.

1.1.1 The Ideal Spiking Neuron
A typical neuron can be divided into three functionally distinct parts, called dendrites, soma, and axon; see Fig. 1.2. Roughly speaking, the dendrites play the role of the ‘input device’ that collects signals from other neurons and transmits them to the soma. The soma is the ‘central processing unit’ that performs an important non-linear processing step: If the total input arriving at the soma exceeds a certain threshold, then an output signal is generated. The output signal is taken over by the ‘output device’, the axon, which delivers the signal to other neurons.
The junction between two neurons is called a synapse. Let us suppose that a neuron sends a signal across a synapse. It is common to refer to the sending neuron as the presynaptic cell and to the receiving neuron as the postsynaptic cell. A single neuron in vertebrate cortex often connects to more than postsynaptic neurons. Many of its axonal branches end in the direct neighborhood of the neuron, but the axon can also stretch over several centimeters so as to reach neurons in other areas of the brain.
| A | B | |
|---|---|---|

|
 |
1.1.2 Spike Trains
The neuronal signals consist of short electrical pulses and can be observed by placing a fine electrode either on the soma or close to the soma or axon of a neuron; see Fig. 1.2. The pulses, so-called action potentials or spikes, have an amplitude of about 100 mV and typically a duration of 1-2 ms. The form of the pulse does not change as the action potential propagates along the axon. A chain of action potentials emitted by a single neuron is called a spike train – a sequence of stereotyped events which occur at regular or irregular intervals; see Fig. 1.3. Since isolated spikes of a given neuron look alike, the form of the action potential does not carry any information. Rather, it is the number and the timing of spikes which matter. The action potential is the elementary unit of signal transmission.
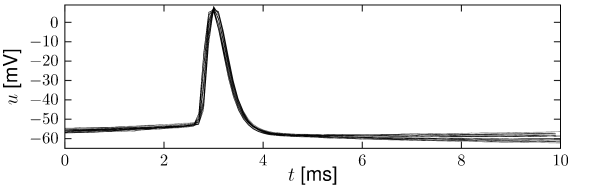
Action potentials in a spike train are usually well separated. Even with very strong input, it is impossible to excite a second spike during or immediately after a first one. The minimal distance between two spikes defines the absolute refractory period of the neuron. The absolute refractory period is followed by a phase of relative refractoriness where it is difficult, but not impossible to excite an action potential.
1.1.3 Synapses
The site where the axon of a presynaptic neuron makes contact with the dendrite (or soma) of a postsynaptic cell is the synapse. The most common type of synapse in the vertebrate brain is a chemical synapse. At a chemical synapse, the axon terminal comes very close to the postsynaptic neuron, leaving only a tiny gap between pre- and postsynaptic cell membrane. This is called the synaptic cleft. When an action potential arrives at a synapse, it triggers a complex chain of bio-chemical processing steps that lead to a release of neurotransmitter from the presynaptic terminal into the synaptic cleft. As soon as transmitter molecules have reached the postsynaptic side, they will be detected by specialized receptors in the postsynaptic cell membrane and lead (either directly or via a biochemical signaling chain) to an opening of specific channels causing ions from the extracellular fluid to flow into the cell. The ion influx, in turn, changes the membrane potential at the postsynaptic site so that, in the end, the chemical signal is translated into an electrical response. The voltage response of the postsynaptic neuron to a presynaptic spike is called the postsynaptic potential.
Apart from chemical synapses neurons can also be coupled by electrical synapses, sometimes called gap junctions. Specialized membrane proteins make a direct electrical connection between the two neurons. Not much is known about the functional aspects of gap junctions, but they are thought to be involved in the synchronization of neurons.
1.1.4 Neurons are part of a big system
Neurons are embedded in a network of billions of other neurons and glial cell that make up the brain tissue. The brain is organized in different regions and areas. The cortex can be thought of as a thin but extended sheet of neurons, folded over other brain structures. Some cortical areas are mainly involved in processing sensory input, other areas are involved in working memory or motor control.
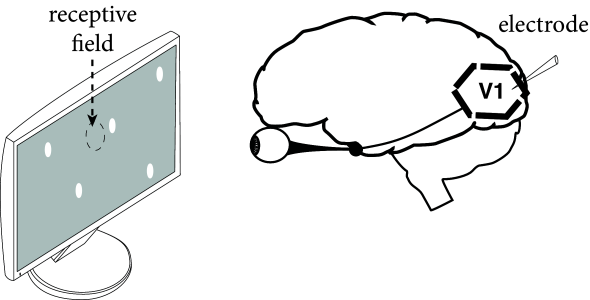
Neurons in sensory cortices can be experimentally characterized by the stimuli to which they exhibit a strong response. For example, neurons in the primary visual cortex respond to dots of lights only within a small region of the visual space. The limited zone where a neuron is sensitive to stimuli is called the neuron’s receptive field (Fig. 1.4).
The receptive field of so-called simple cells in visual cortex is not homogeneous, but has typically two of three elongated subfields. When a light dot falls into one of the positive subfields, the neuron increases its activity, i.e., it emits more spikes than in the absence of a stimulus. Whenever a light dot falls into a negative subfield, it decreases the activity compared to its spontaneous activity in the presence of a gray screen. A spot of light is in fact not the best stimulus. The neuron responds maximally to a moving light bar with an orientation aligned with the elongation of the positive subfield (231).
A large body of the neuroscience literature consists in determining the receptive fields of neurons in sensory cortices. While neurons in visual cortex respond to appropriate visual stimuli, neurons in auditory cortex or somatosensory cortex respond to auditory or tactile stimuli. The concept of receptive field becomes less well defined if one moves away from sensory cortex. For example, in inferotemporal cortex, neurons respond to objects independently of their size and location; in working memory tasks, frontal cortex neurons are active during periods where no stimulus is present at all. In Parts II, III, and IV of this book we touch on aspects of receptive fields and memory of neuronal networks embedded in a big system. For the moment, we return to a simple, idealized neuron.