17.1 Associations and memory
A well-known demonstration of the strong associations, which are deeply embedded in the human brain is given by the following task. The aim is to respond as quickly as possible to three questions. Think of the first answer that comes to mind! Are you ready? Here are the questions: (i) Can you give me an example of a color? – (ii) Can you give me an example of a tool? – (iii) Can you give me an example of a fruit? – For each of these, what was the very first example that came to your mind? Chances are high that your examples are ‘red’ for color and ‘hammer’ for tool. In fact, most humans have particularly strong associations from tool to hammer and from color to red. Regarding fruit, the cultural background plays a more important role (apple, orange, banana), but since the text at the beginning of this chapter mentioned bananas, you probably had a slightly stronger bias toward banana at the moment when you answered the above questions than what you would have had under normal circumstances. This bias through an earlier word or context is a highly significant effect, called ‘priming’ in psychophysics.
Not surprisingly, the word ‘red’ is associated with seeing the color red and vice versa. If you read a list of words that contains names of colors, you are normally fast in doing so and do not experience any particular difficulty. Similarly, you can easily name the color of objects. However, people find it difficult to name the ink color in lists of words that contain entries such as red, green, blue, but are written in colored fonts that are inconsistent with the word (e.g. the word red is written in green font, whereas the word green is written in blue font). In this case responses in the color-naming task are slower compared to naming the color of geometric objects . The measurable difference in reaction time in naming the color of (inconsistent) words compared to the color of objects is called the Stroop effect (500; 320). The association of the color ‘red’ with the word red makes it difficult to name the ink color (e.g., green) in which the word red is written.
In this chapter we mainly focus on association in the sense of completing partial information. Take a look at Fig. 17.1A. Nearly all words are incomplete, but your brain is able to cope with this situation, just as you are able to follow a phone conversation over a noisy line, recognize a noisy image of a handwritten character or associate the picture of an orange with its taste so as retrieve your concept of an orange as a tasty fruit.
| A | B |
|---|---|
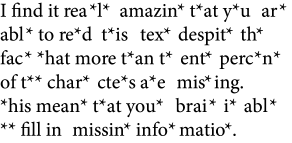
|
 |
17.1.1 Recall, recognition, and partial information
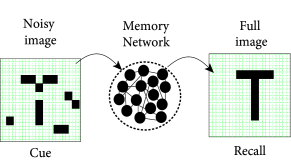
If half of an orange is hidden behind a coffee mug, you can still recognize it as an orange based on the partial information you have. Recognition works because you have seen oranges before and have memorized the concept ‘orange’, including a prototypical image of this fruit. More generally, when you see a noisy image of a known object (e.g., the letter ‘T’) your brain is able to retrieve from memory the prototype version of the object (e.g., an idealized ‘T’). Thus recognizing an object in a noisy environment involves the process of ’memory recall’.
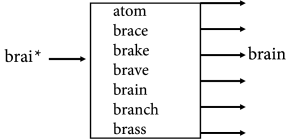
A highly simplified schematic view of memory recall based on partial information is shown in Fig. 17.1B. The input (e.g., an incomplete word) is compared to a list of all possible words. The most likely entry (i.e. the one which is most similar to the input) in the list is given as the output of memory recall.
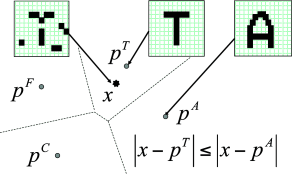
Similarly, noisy images of objects are recognized if the brain finds, among the memorized items, one which is highly similar (Fig. 17.2). Let us call the ‘pure’ noise-free memory item a prototype , where the index labels all different memory items. The prototype can be visualized as a point in some high-dimensional space. A noisy input cue corresponds to another point in the same space. Suppose that we have a similarity measure which enables us to calculate the distance between the input cue and each of the prototypes. A simple method of memory recall is a search algorithm that goes through the list of all available prototypes to find the nearest one. More formally, the output of the recall process is the prototype with
| (17.1) |
which gives rise to a simple geometric picture (Fig. 17.2B).
The aim of this chapter is to replace the explicit algorithmic search for the nearest prototype by the dynamics of interacting neurons. Instead of an explicit algorithm working through a list of stored prototypes, the mere cross-talk of neurons embedded in a large network will find the prototype that corresponds best to the noisy cue – in a highly distributed and automatic fashion, reminiscent of what we believe is happening in the brain (Fig. 17.2A). Brain-style computation implements an implicit algorithm, as we will see in Sections 17.2 and 17.3.
| A | B |
|---|---|
|
 |
17.1.2 Neuronal assemblies
Neural assemblies play a central role in the implicit algorithm for memory retrieval that we will discuss in Section 17.2. Neuronal assemblies (210) are sub-networks of strongly connected neurons that, together, represent an abstract concept. For example, your mental concept of a ‘banana’ containing the mental image of its form, color, taste and texture could be represented by one assembly of strongly connected neurons, while another one might represent your concept of Paris with mental pictures of the Eiffel tower and the Louvre and yet another one your concept of Sydney with its famous opera house.
The assembly as a subgroup of strongly connected neurons has been an influential theoretical notion, introduced by Hebb (210). Do such assemblies exist? The short answer is: We don’t know. Neurons belonging to an assembly do not have to be neighbors but can be widely distributed across one, or even several, brain areas. Experimentally, it is therefore difficult to check for the presence of an assembly as a group of neurons. However, Quiroga et al. (411) found individual neurons in human patients that code for abstract mental concepts such as the Sydney opera house. These patients suffer from severe treatment-resistant epilepsy which makes a surgical intervention necessary. In order to precisely locate the focus of the epilepsy in relation to important brain areas (such as those for speech or motor control), electrophysiological recordings are made while the patient performs various tasks. In contrast to neurons in visual cortex which respond in the presence of an appropriate visual stimulus, single neurons in the medial temporal lobe of human cortex (in particular, in the hippocampus) do not respond to a specific stimulus, but to a much broader set of stimuli that are linked to the same mental concept. For example, the written word ‘Sydney’ and a picture of the opera house in Sydney both cause a response of the same neuron (Fig. 17.3), which we can therefore interpret as one of the neurons belonging to the assembly of neurons encoding the mental concept ‘Sydney’.
| A | B |
|---|---|
|
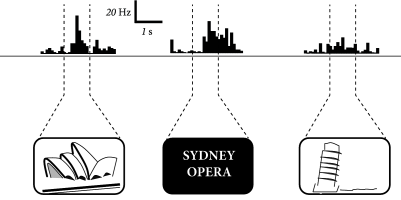 |
Three aspects are worth emphasizing. First, it is unlikely that the neuron responding to the Sydney opera house is the only one to do so. Therefore, we should not think of a single neuron as representing a concept or memory item, but rather a group of neurons. The idea that a single neuron represents one concept is sometimes called the ‘grandmother cell’ code: if the cell coding for grandmother were to die in our brain, our memory of grandmother would disappear as well. At the current stage of research, neural codes based on groups of cells are a more likely code than a grandmother cell code.
Second, the same neuron participates in several assemblies. In the recording sessions of Quiroga et al. where a large collection of pictures of famous individuals and landmarks were used, each unit showed strong responses to about 3 percent of the stimuli (411).
Third, some, but not all, of the neurons showed prolonged responses that persisted after the end of stimulus presentation. This could potentially indicate that a memory item is retrieved and kept in the brain even after the stimulus has disappeared. All three aspects play a role for the memory model discussed in Section 17.2.
17.1.3 Working memory and delayed matching-to-sample tasks
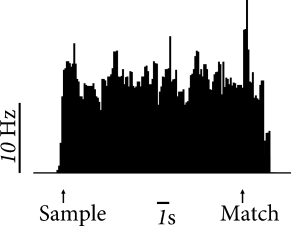
In contrast to long-term memory, items in working memory do not have to be kept for a lifetime. For example, humans use their working memory when they write down a phone number that they just received or search in the supermarket for items on their shopping list. Neural activity during working memory tasks has been recorded in monkeys, in particular in prefrontal and inferotemporal cortex (349; 164; 342). In a delayed matching-to-sample task, a monkey has to indicate whether a second stimulus is, or is not, identical to a first stimulus received one or several seconds earlier.
| A | B |
|---|---|
|
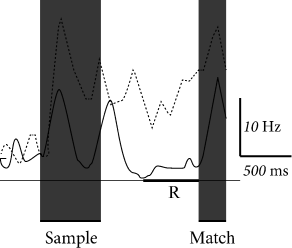 |
In order to correctly perform the task, the monkey has to remember the sample stimulus during the delay period where no stimulation is given. Some neurons in prefrontal cortex show sustained activity during the delay period (Fig. 17.4A). This has been interpreted as a neural signature of working memory. During the delay period, the time course of neural activity varies widely between different objects for one neuron (Fig. 17.4B). and across a population of neurons (412) which indicates that simple models such as the ones discussed in this chapter do not explain all aspects of working memory.