14.3 Linear response to time-dependent input
We consider a homogeneous population of independent neurons. All neurons receive the same time-dependent input current which varies about the mean . For constant input the population would fire at an activity which we can derive from the neuronal gain function. We require that the variations of the input
| (14.41) |
are small enough for the population activity to stay close to the value
| (14.42) |
with .
In that case, we may expand the right-hand side of the population equation into a Taylor series about to linear order in . In this section, we want to show that for spiking neuron models (either integrate-and-fire or SRM neurons) the linearized population equation can be written in the form
| (14.43) |
where is the interval distribution for constant input , is a real-valued function that plays the role of an integral kernel, and
| (14.44) |
is the input potential generated by the time-dependent part of the input current. The first term of the right-hand side of Eq. (14.43) takes into account that previous perturbations with have an after-effect one inter-spike interval later. The second term describes the immediate response to a change in the input potential. If we want to understand the response of the population to an input current , we need to know the characteristics of the kernel . The main task of this section is therefore the calculation of to be performed in Section 14.3.1.
The linearization of the integral equation (183) is analogous to the linearization of the membrane potential density equations (78) that was presented in Chapter 13. In order to arrive at the standard formula for the linear response,
| (14.45) |
we insert Eq. (14.44) into Eq. (14.43) and take the Fourier transform. For we find
| (14.46) |
Hats denote transformed quantities, i.e., is the Fourier transform of the response kernel; is the Fourier transform of the interval distribution ; and is the transform of the kernel . Note that for we have and since and are constant.
The frequency dependent gain describes the linear response of the population activity to a periodic input current . The linear response filter in the time-domain is found by inverse Fourier transform
| (14.47) |
is the mean rate for constant drive . The filter plays an important role for the analysis of the stability of the stationary state in recurrent networks (Section 14.2).
Example: Leaky integrate-and-fire with escape noise
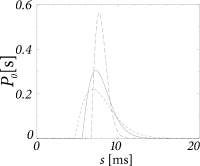
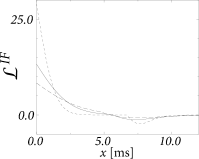
The frequency-dependent gain depends on the filter which in turns depends on the width of the interval distribution (Fig. 14.9B and A, respectively). In Fig. 14.9C we have plotted the signal gain for integrate-and-fire neurons with escape noise at different noise levels. At low noise, the signal gain exhibits resonances at the frequency that corresponds to the inverse of the mean interval and multiples thereof. Increasing the noise level, however, lowers the signal gain of the system. For high noise (long-dashed line in Fig. 14.9C) the signal gain at 1000 Hz is ten times lower than the gain at zero frequency. The cut-off frequency depends on the noise level. The gain at zero frequency corresponds to the slope of the gain function and changes with the level of noise.
| A | B | C |
|---|---|---|
|
|
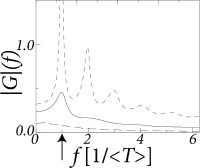 |
14.3.1 Derivation of the linear response filter (*)
In order to derive the linearized response of the population activity to a change in the input we start from the conservation law,
| (14.48) |
cf. Eq. (14.8). As we have seen in Section 14.1 the population equation (14.5) can be obtained by taking the derivative of Eq. (14.8) with respect to , i.e.,
| (14.49) |
For constant input , the population activity has a constant value . We consider a small perturbation of the stationary state, , that is caused by a small change in the input current, . The time-dependent input generates a total postsynaptic potential, where is the postsynaptic potential for constant input and
| (14.50) |
is the change of the postsynaptic potential generated by . Note that we keep the notation general and include a dependence upon the last firing time . For leaky integrate-and-fire neurons, we set whereas for SRM neurons we set . We expand Eq. (14.49) to linear order in and and find
| (14.51) |
We have used the notation for the survivor function of the asynchronous firing state. To take the derivative of the first term in Eq. (14.51) we use and . This yields
| (14.52) |
We note that the first term on the right-hand side of Eq. (14.52) has the same form as the population integral equation (14.5), except that is the interval distribution in the stationary state of asynchronous firing.
To make some progress in the treatment of the second term on the right-hand side of Eq. (14.52), we now restrict the choice of neuron model and focus on either SRM or integrate-and-fire neurons.
(i) For SRM neurons, we may drop the dependence of the potential and set where is the input potential caused by the time-dependent current ; compare Eqs. (14.44) and (14.50). This allows us to pull the variable in front of the integral over and write Eq. (14.52) in the form
| (14.53) |
with a kernel
| (14.54) |
(ii) For leaky integrate-and-fire neurons we set , because of the reinitialization of the membrane potential after the reset (183). After some rearrangements of the terms, Eq. (14.52) becomes identical to Eq. (14.53) with a kernel
| (14.55) |
Let us discuss Eq. (14.53). The first term on the right-hand side of Eq. (14.53) is of the same form as the dynamic equation (14.5) and describes how perturbations in the past influence the present activity . The second term gives an additional contribution which is proportional to the derivative of a filtered version of the potential .
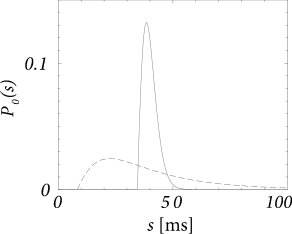
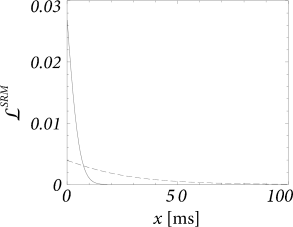
We see from Fig. 14.10 that the width of the kernel depends on the noise level. For low noise, it is significantly sharper than for high noise.
| A | B |
|---|---|
|
 |
Example: The kernel for escape noise (*)
In the escape noise model, the survivor function is given by
| (14.56) |
where is the instantaneous escape rate across the noisy threshold; cf. Chapter 7. We write . Taking the derivative with respect to yields
| (14.57) |
where and . For SRM-neurons, we have and , independent of . The kernel is therefore
| (14.58) |
Example: Absolute refractoriness (*)
Absolute refractoriness is defined by a refractory kernel for and zero otherwise. We take an arbitrary escape rate . The only condition on is that the escape rate goes rapidly to zero for voltages far below threshold: .
This yields and hence
| (14.59) |
The survivor function is unity for and decays as for . Integration of Eq. (14.58) yields
| (14.60) |
As we have seen in Section 14.1, absolute refractoriness leads to the Wilson-Cowan integral equation (14.10). Thus defined in (14.60) is the kernel relating to Eq. (14.10). It could have been derived directly from the linearization of the Wilson-Cowan integral equation (see Exercises). We note that it is a low-pass filter with cut-off frequency , which depends on the input potential .