16.1 Perceptual Decision Making
Many perceptual phenomena can be formulated as a problem of decision making. In a typical experiment of visual psychophysics, a subject observes a short flash of three vertical black bars on a gray background (Fig. 16.1B). Is the middle bar shifted to the left or to the right compared to a symmetric arrangement of the three bars where it is exactly in the center? If the shift is very small, or if the bars are presented with low contrast on a noisy screen, the question is difficult to answer. The subject who holds a button in each hand, indicates his decision (left or right) by pressing the corresponding button. In other words, he reports his perception as a decision.
In the following, we focus on an experimental paradigm with visual random dot motion stimuli used for the study of perceptual decision making in monkeys (454; 443; 191). The stimulus consists of a random pattern of moving dots, where most, but not necessarily all, of the dots move coherently in the same direction; cf. Fig. 16.2. Typically, two different directions of motion are used, e.g., upward or downward. The monkey has been trained to indicate the perceived motion direction by saccadic eye movements to one of two targets cf. Fig. 16.2B.
Note that, in contrast to the examples given at the beginning of this chapter, problems of perceptual decision making typically have no direct monetary value or risk associated with it. Normally we do not care whether a bar is shifted to the left or to the right, or whether dots move upward or downward. Nevertheless, a correct perceptual decision might be life-saving if a moving stripe pattern in the bush is correctly recognized as an approaching tiger as opposed to a movement of the leaves in the wind.
16.1.1 Perception of motion
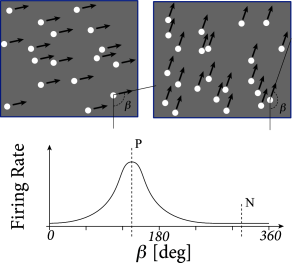
Neurons in the middle temporal visual area (MT, also called V5) are activated by large-scale motion stimuli. The receptive field of an MT neuron, i.e., the region of visual space it is sensitive to motion stimuli, is considerably larger than that of a neuron in primary visual cortex; cf. Chapter 12. Different neurons in MT respond to different directions of motion, but just as in other parts of visual cortex, area MT has a columnar structure so that clusters of neighboring neurons share receptive fields with a similar preferred direction of motion (14).
At the beginning of a typical recording session with an extracellular electrode in MT (454), the location of the receptive field and the preferred direction of motion of a single neuron or cluster of neighboring neurons is determined by varying the movement angle and the location of the random dot stimulus. Once the receptive properties of the local MT neurons have been determined, only two different classes of stimuli are used, i.e., dots moving coherently in the preferred direction of the recorded neuron, and dots moving coherently in the opposite direction.
After each presentation of a random dot motion pattern, two targets are switched on, one at a location in direction of stimulus motion, the other one on the opposite side. The monkey is trained to indicate the movement direction of the stimulus by a saccadic eye movement to the corresponding target. After training, the perceptual decision between a dot movement in the cell’s preferred direction (P) or the null direction (N) is reliably performed by the monkey, if a noise-free stimulus is used where all dots move in the same direction. However, the task becomes more difficult, if only a small fraction of dots move coherently in one of the two directions while the rest of the dots moves in a random direction. The behavioral performance can be assessed with the psychometric function which represents the percentage of saccades to the target P as a function of coherence, where coherence indicates the fraction of coherently moving dots (Fig. 16.2B).
| A | B |
|---|---|
|
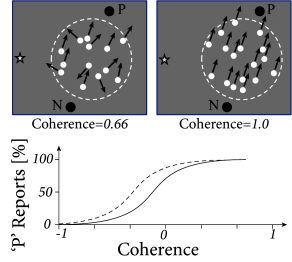 |
An electrode in MT cannot only be used to record neural activity, but also to stimulate a cluster of neurons in the neighborhood of the electrode. Since neighboring neurons have similar preferred directions of motion, current injection into the electrode can bias the perception of the monkey in favor of the neurons’ preferred direction, even if the random dot pattern has no or only a small amount of coherence (Fig. 16.2B). This indicates that the perceptual decision of the monkey relies on the motion information represented in the activity of MT neurons (454).
While the monkey’s perceptual decision is influenced by the manipulation of MT neurons, this result does not imply that the decision itself is made in MT. It is likely to be made at a later stage, in an area that uses the information of MT neurons.
16.1.2 Where is the decision taken?
The short answer is: we do not know. However, an interesting observation has been made in the Lateral Intra-parietal Area (LIP) during experiments of perceptual decision making with moving random dot stimuli (443).
Before discussing the experiment, we need to present a few facts about the properties of LIP neurons. Area LIP is located in the visual processing stream between primary visual cortex and the Frontal Eye Field region involved in control of saccadic eye movements. Neurons in area LIP respond during the preparation of saccadic eye movements. Different neurons in LIP have different receptive field. The location of the receptive field corresponds to a potential target region of eye movements. In other words, a LIP neuron responds just before a saccadic eye movement into its receptive field occurs.
As in the previous subsection, monkeys in the experiment of Roitman and Shadlen are trained to indicate the direction of a moving dot pattern by saccadic eye movements to one of two visual targets. The first target is located in the receptive field of a LIP neuron. Therefore, the recorded neuron is expected to respond whenever the monkey prepares a movement to the first target. The second target is located in the opposite direction. The task is designed such that a random dot stimulus moving in direction of the first target indicates that the monkey should make an eye movement toward it; the correct response to a stimulus moving in the opposite direction is a saccade to the second target (Fig. 16.3A). The difficulty of the task can be varied by changing the fraction of coherently moving dots. The behavioral reaction time of the monkey was measured as a function of stimulus coherence. At the same time, the activity of neurons in LIP was recorded.
| A | B |
|---|---|

|
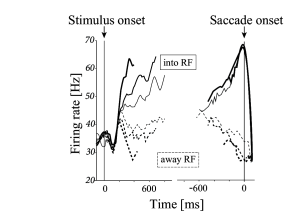 |
Roitman and Shadlen found that, during the presentation of the moving dot stimulus, the activity of LIP neurons increased. The rate of increase after stimulus onset was higher for stimuli with a large fraction of coherent points than for stimuli with little or no coherence. Importantly, when the responses were averaged and aligned to the onset of the saccade, LIP neurons always reached the same level of activity just before a saccade into their receptive field (Fig. 16.3B).
These findings are consistent with the idea that the decision to perform a saccade occurs at the moment when LIP neurons reach a threshold value. For stimuli with a high degree of coherence, the activity increases more rapidly, the threshold is reached earlier, and reaction times are shorter than for stimuli with low degree of coherence. Therefore, the paper of Roitman and Shadlen suggests that ‘a threshold level of LIP activity appears to mark the completion of the decision process’ (443).